研究内容
研究概要
研究目的
人口の増加、耕地の減少、資源の枯渇、さらには経済のグローバル化により、食糧・植物バイオマス資源の必要量は急速に増加しています。これに対応するためには、不良な土壌での省資源型の生産体系を確立することが必要と考えられています。不良土壌とは、酸性土壌、塩類集積土壌、重金属集積土壌をさす言葉ですが、実に全世界の"農耕地""林地"として使用できる土壌の70%は不良土壌に分類されます。これらの土壌では、植物の生産性が低く、それを補うためには、資源・エネルギーを投入することが必要になります。私たちは、このような環境でも、少ない資源・エネルギー投入で生産を行える"耐性植物"を作出することを目的として、植物個体レベルから分子・ゲノムレベルでの研究を進めています。

典型的な酸性土壌
南アフリカで撮影した酸性土壌の畑です。写真の奥では1haあたり10tの石灰を投入して酸性を中和し、ヨーロッパの人たちが食べる食料を生産しています。酸性土壌では、リン酸肥料も多量に必要となります。手前側の無石灰区では、植物は根がはれず乾燥期に枯死します。私たちが利用する食糧・バイオマスの生産でも、同じことが起きています。

酸性土壌とは?
温帯から熱帯の多雨地域(年間降水量が多い地域)では、土壌中の塩基(CaやMgなど)が雨水の浸透と共に失われ土壌は酸性化します。この土壌では、植物に対して毒性が強いアルミニウムが可溶化して根の伸長が阻害され、植物は旱魃抵抗性を失い、養分吸収能力も低くなります。酸性土壌では、養分欠乏も収量低下の原因となります。写真は、典型的なマグネシウム欠乏を呈しており、肥料を与えることが経済的に困難な場合、生産性は低下します。

酸性土壌耐性メカニズムの解明に向けて
酸性土壌耐性には、種間差・品種間差が存在します。小麦の耐性(右)・感受性(左)品種を石灰投入量を変えた酸性土壌で栽培すると、耐性品種は石灰投入量が少なくても生育できることがわかります。その生理機構を遺伝子のレベルで解明することを"分子生理学"と呼び、生化学・遺伝学・分子生物学・ゲノム生物学などの手法を用いて研究します。私たちのグループでは、耐性と感受性の植物種や品種を比べることで、耐性原因遺伝子の特定と耐性メカニズムの解明を目指します。

シロイヌナズナの自然変異
シロイヌナズナは世界中に広く分布しており、生息地間でゲノム配列や環境ストレス耐性が異なります(右写真)。そのような遺伝的多様性を示す集団において、ゲノムワイド関連解析(GWAS)やトランスクリプトーム解析により、遺伝情報とストレス耐性との関連を調べます。私たちは、それらを通じてミネラルストレス対する耐性/感受性の遺伝的要因の探索を行い、原因遺伝子の同定と機能解析を進めます。ストレス耐性多様性の分子メカニズムを解明することで、分子育種に利用できる情報基盤の構築に貢献したいと考えます。
主な研究内容
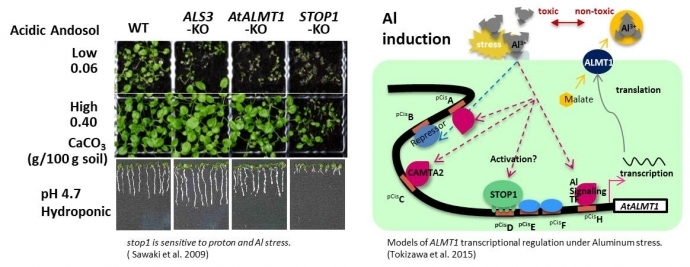
リンゴ酸トランスポーターAtALMT1遺伝子の発現制御に関する研究
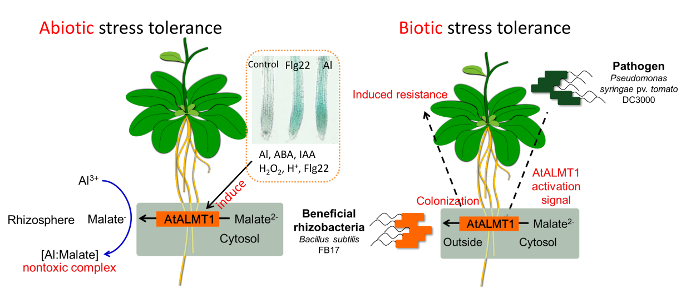
酸性土壌では、細胞毒性が高いアルミニウム(Al)が可溶化します。これにより、根は伸長できなくなります。また、アルミニウムは施肥の3大要素であるリンと難溶性のリン酸塩を作り、リン酸欠乏をもたらします。1)Al耐性には、根へのAlの到達を防ぐ"Al排除機構"と、内部で生じる問題を解決・修復する"内部Al耐性機構"があります。最も重要な耐性機構には有機酸が関係しています。クエン酸、リンゴ酸などは、Alと錯体(キレート)を形成して体内・土壌中でAlを無毒化します。シロイヌナズナでは、根圏へのリンゴ酸排出はリンゴ酸トランスポーターAtALMT1が担っています(Kobayashi et al. 2007, Plant Physiol.)。2)低リン酸耐性には、土壌中で難溶化しているリン酸Alを利用する能力が関係しています。根からクエン酸を放出すれば、リン酸を溶かして利用することが可能になります。つまり、根からクエン酸・リンゴ酸を放出する能力は、酸性土壌で良好に生育するために有利な形質といえます(Koyama et al. 2000, Plant Cell Physiol.)。3)AtALMT1遺伝子の転写は、アルミニウムイオンだけでなく、様々なストレス因子や植物ホルモンに応答して誘導され(Kobayashi et al. 2013, Plant Physiol.)、植物免疫の活性化にもAtALMT1を介した有用菌の根圏への誘引が関係しています(Kobayashi et al. 2013, Plant Signal Behav.) 。私たちは、その転写活性化の分子機構を解明するため、プロモーターや発現調節因子の特定に取り組んでいます。
STOP1転写因子の制御機構に関する研究
シロイヌナズナ酸感受性変異株の原因遺伝子として単離したSTOP1(Sensitive TO Proton rhizotoxicity 1)遺伝子は、C2H2 型ジンクフィンガータンパク質をコードしており、酸耐性遺伝子だけでなく、多くのAl耐性遺伝子の発現を制御する転写調節因子です(Iuchi et al. 2007, PNAS. Sawaki et al. 2009, Plant Physiol. Kobayashi et al. 2013, Mol Plant, Tokizawa et al. 2015, Plant Physiol.)。また、STOP1による転写制御機構は、多くの陸上植物種で保存され機能していることが分かってきました(Ohyama et al. 2013, Plant Physiol. Sawaki et al. 2014, Plant Sci. Daspute et al. 2018, Planta) 。私たちは、STOP1が制御する遺伝子の役割と発現制御の仕組みを解明し、植物の環境ストレス耐性におけるSTOP1システムを明らかにすることで実用植物への応用展開を目指します。